
Ancient DNA from Sudan
11 Saturday Mar 2023
Tags
A-Group, Afro-Asiatic, ancient DNA, C-Group, Christian Nubia, Kerma, Meroe, Northeast Africa, Savanna Pastoral Neolithic
Linguists often hypothesize that the Sudan area in Northeast Africa may have been the urheimat or "original homeland" of the Afro-Asiatic (Hamitic-Semitic) language family. Certain old cultures in the vicinity — namely, the A-Group, C-Group, Kerma and Meroitic civilizations — also have close ties with that of ancient Egypt and the Cushitic-associated Savanna Pastoral Neolithic. As such, the biological affinities of early populations from this region are of considerable interest to scientists.
Ancient DNA (aDNA) analysis has become increasingly common in Europe and parts of Asia. This is due to the generally temperate climate in these geographical areas, which in turn facilitates the molecular preservation of DNA extracted from ancient human specimens. Conversely, much of the northern half of Africa as well as the Middle East falls under a hot desert climate (BWh) in the global Köppen climate classification:
 As a result, the typically hot and arid environmental conditions in these regions have until recently made the collection of ancient DNA difficult.
As a result, the typically hot and arid environmental conditions in these regions have until recently made the collection of ancient DNA difficult.
Neolithic and Meroitic, Post-Meroitic & Christian periods
In 2009, Hisham Yousif and Muntaser Eltayeb with the University of Khartoum managed to successfully extract ancient DNA from bone samples stored at the Sudan National Museum. The specimens dated from the Neolithic, Meroitic, Post-Meroitic and Christian periods, with the Neolithic specimens belonging to the Kadruka culture:
 The researchers observed that the Neolithic Kadruka samples generally belonged to different paternal lineages than the later Meroitic, Post-Meroitic and Christian period samples. Most of the Kadruka individuals were haplogroup A-M13 bearers, whereas the Meroitic, Post-Meroitic and Christian era samples were instead mainly haplogroup DE (haplogroup YAP, which includes the D-M174 and E-M96 clades) and haplogroup F-M89 carriers:
The researchers observed that the Neolithic Kadruka samples generally belonged to different paternal lineages than the later Meroitic, Post-Meroitic and Christian period samples. Most of the Kadruka individuals were haplogroup A-M13 bearers, whereas the Meroitic, Post-Meroitic and Christian era samples were instead mainly haplogroup DE (haplogroup YAP, which includes the D-M174 and E-M96 clades) and haplogroup F-M89 carriers:
Haplogroups A-M13 was found at high frequencies among Neolithic samples. Haplogroup F-M89 and YAP appeared to be more frequent among Meroitic, Post-Meroitic and Christian periods. Haplogroup B-M60 was not observed in the sample analyze.
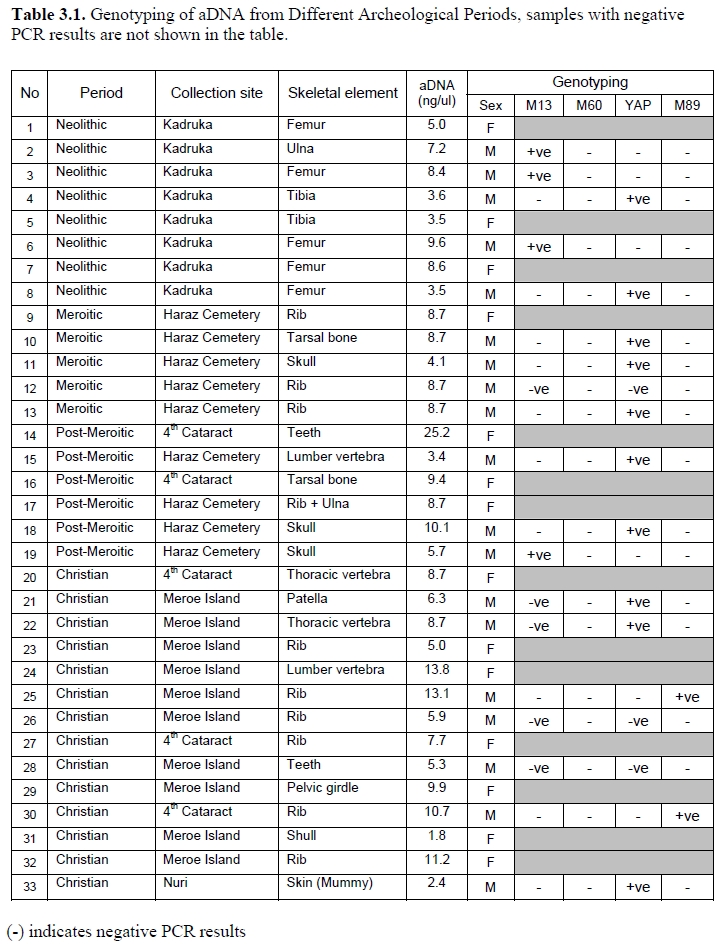 Since haplogroup A is today the most common paternal lineage among Nilotic groups in the Nile Valley, the researchers propose that the Neolithic Kadruka population was likely ancestral to present-day Nilotes residing in the area. By contrast, the higher haplogroup E and F frequencies found among the Meroitic, Post-Meroitic (X-Group) and Christian era populations prompted the scientists to assert that these other early Sudanese groups instead had closer ties with communities ancestral to the modern Afro-Asiatic-speaking (Beja, Sudanese Arabs) and Nubian-speaking populations:
Since haplogroup A is today the most common paternal lineage among Nilotic groups in the Nile Valley, the researchers propose that the Neolithic Kadruka population was likely ancestral to present-day Nilotes residing in the area. By contrast, the higher haplogroup E and F frequencies found among the Meroitic, Post-Meroitic (X-Group) and Christian era populations prompted the scientists to assert that these other early Sudanese groups instead had closer ties with communities ancestral to the modern Afro-Asiatic-speaking (Beja, Sudanese Arabs) and Nubian-speaking populations:
In Y-chromosome terms this mean in simplest terms introgression of the YAP insertion (haplogroups E and D), and Eurasian Haplogroups which are defined by F-M89 against a background of haplogroup A-M13. The data analysis of the extant Y-chromosomes suggests that the bulk of genetic diversity appears to be a consequence of recent migrations and demographic events mainly from Asia and Europe, evident in a higher migration rate for speakers of Afro-Asiatic as compared to the Nilo-Saharan family of languages, and a generally higher effective population size for the former.

An eggshell pottery vase belonging to the A-Group culture (3400-2400 BCE) of Nubia.
Although specimens belonging to the A-Group culture of Lower Nubia (which postdates the Neolithic Kadruka culture of Upper Nubia) were not examined, existing skeletal analysis of these individuals suggests that they too were ancestral to the modern Afro-Asiatic-speaking and Nubian groups. This is because the A-Group specimens share close osteological ties with the latter populations, as well as with ancient and contemporary Egyptian samples. Conversely, the A-Group people are markedly unlike the modern Nilotic groups. This indicates that the A-Group culture bearers in all likelihood have different ancestral origins from the Nilo-Saharan-speaking communities (cf. Billy (1981b)).

Inner coffins of the ancient Egyptian aristocrats Nakht-Ankh (right) and Khnum-Nakht (left), dating from the Middle Kingdom (c. 1985-1773 BCE). Forensic analysis of the bodies of these Two Brothers revealed profound anatomical differences, which indicates that the siblings likely had been placed in the wrong coffins. Nakht-Ankh was found to have had a "Caucasoid" skull of a delicate character, an orthognathous craniofacial profile, a slender build, an olive complexion (based on examination of residual skin tissue), and soft-textured dark brown hair, whereas Khnum-Nakht had a "Negroid" skull of a robust character, a considerably prognathous craniofacial profile, and a dark complexion (cf. Murray (1910); David (2007); Riggs (2014); Forshaw (2019)). Along with some hieroglyphic evidence, this marked physical divergence further suggested to researchers that the brothers shared a mother but had different fathers (Drosou et al. (2018); Forshaw (2019)). Ancient DNA analysis confirmed these suspicions: the men bore different paternal markers but belonged to the same maternal lineage, the M1a1 haplogroup, which is a signature mtDNA clade of the Afro-Asiatic-speaking populations inhabiting the Nile Valley and Horn of Africa (Drosou et al. (2018)). While Khnum-Nakht's specific Y-DNA haplogroup could not be identified, Nakht-Ankh was, after some initial uncertainty, successfully assigned to the H2m clade (cf. HaploTree; Open Genomes). The H2 patrilineage has been tentatively detected in Christian-era Nubian individuals (Yousif and Eltayeb (2009); see discussion below), consistent with the proposed ancestral ties between the "red" Nubians of the ancient Egyptian monuments and the Egyptians themselves.
(*N.B. After the publication of Yousif and Eltayeb (2009)'s study, it was recognized that the M282 mutation, which at the time was classified under F-M89, had been misclassified. M282 was subsequently reassigned to its proper lineage, the paternal haplogroup H; specifically, as the defining mutation of the H2 sublineage. In the archaeogenetic record, H2 mainly has been found among Neolithic European specimens (e.g. Cassidy et al. (2020)). The oldest example of this subhaplogroup in Africa has been detected in an ancient Egyptian individual, the Middle Kingdom nobleman Nakht-Ankh (HaploTree; Open Genomes). Ergo, the instances of F-M89 which Yousif and Eltayeb (2009) identified among their Christian-era Nubian samples might indeed have been inherited from Afro-Asiatic-speaking ancestors.)
Mesolithic, Neolithic and MXCH (Meroitic, Post-Meroitic & Christian periods)
In 2014, a genetic research team led by Kendra Sirak of Emory University's Department of Anthropology managed to successfully extract endogenous DNA from other ancient individuals from Sudan. The analyzed specimens consisted of six samples spanning from the Mesolithic through to the Meroitic, Post-Meroitic/X-Group and Christian periods (MXCH) in Wadi Halfa, Lower Nubia/northern Sudan:
 As of August 2015, the researchers have only published the aDNA results of one of these ancient specimens, a Christian-period infant dubbed NUB04b in the sample set:
As of August 2015, the researchers have only published the aDNA results of one of these ancient specimens, a Christian-period infant dubbed NUB04b in the sample set:
 NUB04b was found to belong to the mtDNA haplogroup L5 (formerly known as L1e), a small maternal clade that today is most common in East Africa:
NUB04b was found to belong to the mtDNA haplogroup L5 (formerly known as L1e), a small maternal clade that today is most common in East Africa:
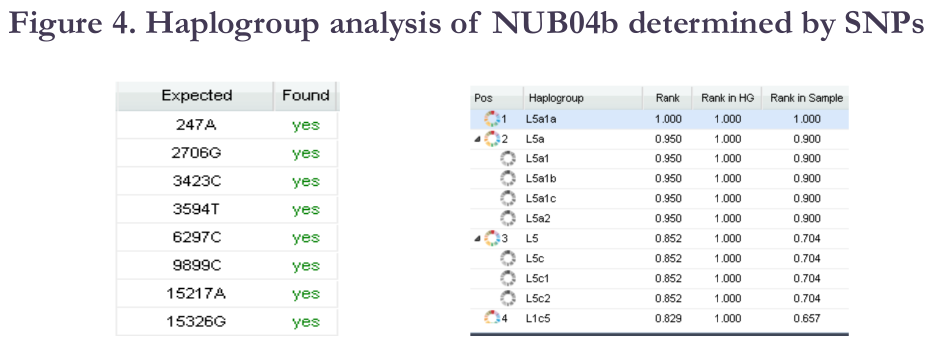 To gauge NUB04b's autosomal DNA affinities, the researchers also ran a principle component analysis, which compared 300 of the specimen's single nucleotide polymorphisms (SNPs) spatially against those of various modern individuals from the Omni 2.5 dataset. For reasons I shall explain in a future post, such biogeographical testing/admixture testing, including SNP genotyping and whole genome analysis, is extremely flawed with regard to present-day populations alone (this in large part has to do with the difference between the genealogical tree and the genetic tree). It can, however, be informative for ancient individuals when they are compared against modern individuals. Unfortunately, NUB04b's SNP genotyping was apparently compromised by the presence of some extraneous DNA:
To gauge NUB04b's autosomal DNA affinities, the researchers also ran a principle component analysis, which compared 300 of the specimen's single nucleotide polymorphisms (SNPs) spatially against those of various modern individuals from the Omni 2.5 dataset. For reasons I shall explain in a future post, such biogeographical testing/admixture testing, including SNP genotyping and whole genome analysis, is extremely flawed with regard to present-day populations alone (this in large part has to do with the difference between the genealogical tree and the genetic tree). It can, however, be informative for ancient individuals when they are compared against modern individuals. Unfortunately, NUB04b's SNP genotyping was apparently compromised by the presence of some extraneous DNA:
The sample is found to be located between European and African populations, suggesting the presence of some European contaminants extracted together with endogenous DNA.
So what can be gleaned from this ancient DNA analysis? For starters, certain mtDNA haplogroups that today are primarily concentrated below (or above) the Sahara may not always have been. Such appears to be the case with the L5 clade to which the NUB04b specimen apparently belongs. More importantly, these lineages may originally have had quite different population affinities than their present-day distribution would suggest.
As the earliest ancient specimen so far found to carry haplogroup L5, it would be interesting to know just what are NUB04b's general phenotypic affinities. To this end, the researchers provide a graphic illustrating the marked difference in skull form between the later Meroitic, Post-Meroitic/X-Group and Christian (MXCH) period inhabitants of Wadi Halfa and the earlier Mesolithic period dwellers:
 The Mesolithic period inhabitants were clearly a more robust people, with a rugged osteological framework. Comparative craniofacial and anthropometric analyses of these ancient specimens versus later MXCH samples, modern Nubians and ancient and modern Egyptian samples have been conducted by various anthropologists. These analyses have consistently grouped the Mesolithic Wadi Halfa crania with those of present-day "Negroid" populations, while the MXCH and modern Nubian samples have instead typically clustered with Egyptians and other surrounding populations with a generally "Caucasoid" craniofacial pattern.
The Mesolithic period inhabitants were clearly a more robust people, with a rugged osteological framework. Comparative craniofacial and anthropometric analyses of these ancient specimens versus later MXCH samples, modern Nubians and ancient and modern Egyptian samples have been conducted by various anthropologists. These analyses have consistently grouped the Mesolithic Wadi Halfa crania with those of present-day "Negroid" populations, while the MXCH and modern Nubian samples have instead typically clustered with Egyptians and other surrounding populations with a generally "Caucasoid" craniofacial pattern.





-
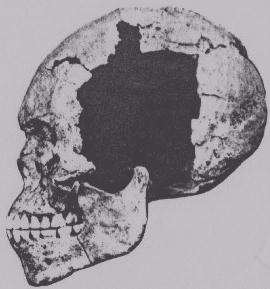
- A Mesolithic skull shown in profile, which was excavated at Wadi Halfa, Nubia. Note the marked prognathism (facial projection). Craniometric analysis of the Mesolithic Wadi Halfa crania indicates that these ancient specimens are most closely related to Niger-Congo-speaking individuals. The Afro-Asiatic-speaking samples instead share affinities with the ancient and modern European samples, Bronze Age Nubian (X-Group) and Medieval Nubian (Christian period) samples, and Indo-European-speaking and Dravidian-speaking Indian samples (see Brace (1993) above).

Three different types of crania excavated in Nubia by Grafton Elliot Smith and other Egyptologists accompanying the Archaeological Survey of Nubia. Left=Skull of an individual of unknown origin; Center=Skull of an ancient Egyptian individual; Right=Skull of a "Negro" individual. Notice how the Egyptian cranium closely resembles the MXCH skull above, in accordance with the archaeogenetic evidence (discussed below) indicating that Egyptian-related peoples inhabited areas of Nubia (Elliot Smith and Jones (1910)).
In April 2016, Sirak and colleagues published an abstract on morphological and genetic analysis, which they conducted on ancient Nubian individuals. The sample set consisted of 150 adults excavated in Upper and Lower Nubia, with the specimens dating from the Mesolithic through to the Christian periods. The researchers observed a major difference in craniofacial form between the Mesolithic and Neolithic individuals, in accordance with their haplogroup and SNP analysis. While the briefing does not specify if the Neolithic samples belonged to the earlier Kadruka culture of Upper Nubia or the ensuing A-Group civilization of Lower Nubia, we can assume from the morphological patterns that they were associated with the latter cultural sphere. All in all, the data further underlines the distinct ancestral origins of the Mesolithic period inhabitants of Nubia versus the later A-Group, C-Group, Kerma, Meroitic, X-Group (Post-Meroitic) and Christian era populations:
The biological history of the occupants of Nubia throughout the Holocene remains a topic of debate; specifically, there is some tendency to assume a discontinuous history of occupation and cultural development based on traditional anthropological and archaeological evidence. Population movements, a shift in subsistence strategies and in-situ evolution have all been used to explain the biological variation observed in Nubian remains, particularly in the cranium. Here we investigate craniofacial and mandibular shape patterns, as well as ancient DNA variation, among populations from Upper and Lower Nubia spanning 12,000 years and reflecting a transition from hunting-gathering to intensive farming.
Our sample includes 150 adult specimens from six archaeological sites along the Nile River in Egypt and Sudan that belong to eight chrono-cultural groups spanning from the Late Mesolithic through the Christian periods. All individuals were digitized with a surface scanner, then 397 and 120 three-dimensional landmarks and semilandmarks respectively were extracted on skulls and mandibles of each specimen. Landmark configurations were subjected to generalized Procrustes analysis, tangent space projection, principal component analysis, discriminant analysis, and MANOVA. Endogenous DNA was extracted and sequenced from a subsample in order to explore variation in Nubian population genetics throughout time and space using informative SNPs and haplogroup determination.
Our results highlight a strong distinction between Mesolithic and the Neolithic samples. Craniofacial patterns underline the importance of gene flow and give some support to the hypothesis of regional continuity among more recent groups, while patterns of mandibular morphology show high correlation with subsistence strategy.

Approximate location of the Kadruka archaeological site, situated north of Dongola and south of Kerma along the Nile River in Upper Nubia (Geographic.org). During the early Neolithic period, Kadruka was inhabited by carriers of the archaic African haplogroup A, a paternal lineage that is common today among Nilotic groups in Sudan (cf. Yousif and Eltayeb (2009)'s ancient DNA analysis). Subsequent population movements in the late Neolithic/Bronze Ages would introduce the first Afro-Asiatic speakers into the area. These new arrivals were instead found to be genetically indistinguishable from the Cushitic settlers of the Early Pastoral Neolithic, supporting the theory that these pastoralists had migrated southward to eastern Africa from a center in the Nile Valley (Wang et al. (2022)).
Wang et al. (2022) sequenced DNA from an ancient individual excavated at Kadruka, Upper Nubia. The specimen is 4000 years old and is thus coeval with the Mota hunter-gatherer from southwestern Ethiopia, as well as with Djehutynakht, the oldest Egyptian individual thus far analyzed (who belongs to the mtDNA haplogroup U5b2b5), and the Afro-Asiatic-speaking Kerma and C-Group populations of Nubia. Unsurprisingly, considering the time period in question, the researchers observed that their Middle Kingdom Kadruka specimen was genetically indistinguishable from the Cushitic settlers of the Early Pastoral Neolithic in the Great Lakes region. This reflects the spread of Afro-Asiatic speakers into Upper Nubia, an area which, as seen above (cf. Yousif and Eltayeb (2009)), was formerly in early Neolithic times principally inhabited by haplogroup A carriers.
Christian period
In 2015, Sirak et al. published another conference paper on ancient DNA from the Sudan region. The bone samples this time were collected from the medieval site of Kulubnarti, located between the Second Cataract and Dal Cataract beside the Nile. Three individuals in total, they all dated from the early Christian era i.e., the historical period when the Alodia/Alwah, Nobatia and Makuria kingdoms were extant:

 The scientists then ran an analysis on the sample that yielded the highest endogenous DNA, KulR17. Like NUB04b from Wadi Halfa, this early Kulubnarti inhabitant also happened to be an infant. 523 of KulR17's SNPs were compared against various global reference samples from the Human Genome Diversity Project dataset. However, there was apparently no DNA contamination this time. The researchers instead remarked that "the authenticity of KulR17 seems to show that even in hot and arid climatic conditions it is possible to extract endogenous DNA from archaeological samples."
The scientists then ran an analysis on the sample that yielded the highest endogenous DNA, KulR17. Like NUB04b from Wadi Halfa, this early Kulubnarti inhabitant also happened to be an infant. 523 of KulR17's SNPs were compared against various global reference samples from the Human Genome Diversity Project dataset. However, there was apparently no DNA contamination this time. The researchers instead remarked that "the authenticity of KulR17 seems to show that even in hot and arid climatic conditions it is possible to extract endogenous DNA from archaeological samples."
KulR17's biogeographical ancestry turned out to be most similar to that of modern populations in the Middle East and environs:
From this analysis, the geographic ancestry of this individual was estimated to be closer to Middle Eastern/and Central and South Asian populations than to any African population.


Medieval illustration of a Nubian king.
This finding is consistent with the Coptic ancestral component that Dobon et al. (2015) observed to be the defining element among Egyptian Copts and other Afro-Asiatic speakers in the Nile Valley and Ethiopia, as well as among many present-day Nubians. Hodgson et al. (2014) found an analogous West Eurasian ancestral component among Afro-Asiatic speakers in the Horn region, with a frequency peak among ethnic Somalis. Since it is unlikely that there was a population replacement among Nubians in the intervening centuries after the medieval Christian period, we can safely assume that the Coptic/Ethio-Somali ancestral component that defines modern Nubians is the same West Eurasian-affiliated ancestral component that defines the KulR17 specimen from Kulubnarti.
Additionally, this aDNA result is in keeping with the aforenoted osteological affinities of MXCH period skeletons from Sudan, as well as medieval iconography of Nubian royalty (see illustration on right). In future posts, we shall see that this finding is also in alignment with the linguistic affiliations of the earlier C-Group and Kerma Afro-Asiatic-speaking cultures, and possibly also that of the Meroitic civilization.
*Update #1

Location of Meroë and other key sites in Nubia (Khalil and Miller (1996)).
Exciting news, folks! It took a few years, but we finally have some ancient DNA analysis on pre-Christian Nubia (better late than never).
In early 2020, Yahia Mehdi Seddik Cherifi of the Laboratoire d'Anthropométrie, du Patrimoine et d'Archéologie and Selma Amrani of the Institute of Archaeology at the University of Algiers released a study on the first mitochondrial DNA extracted from individuals belonging to the X-Group (Post-Meroitic) culture. This civilization, also known as the Ballanean Culture, existed in Upper Nubia between 300-600 CE, arising after the end of the Meroitic Kingdom. The X-Group Nubians were culturally different from the preceding Meroites. They also, apparently, did not speak the Meroitic language. They are thought to have spoken instead the Nobiin or Nubian language, a Nilo-Saharan tongue, which most Nubians today speak.
(*N.B. Nubians are a distinct people from the Nuba/Noba. Nubians today speak Nobiin, a Nilo-Saharan language which they are believed to have adopted. Their Meroitic ancestors appear to have originally spoken an Afro-Asiatic language like the neighboring Egyptians (see discussion below). Nubians inhabit the area between central Sudan and southern Egypt. They are thought to descend from the Egyptian-related inhabitants of ancient Nubia (the "red Nubians") because they share close physical, genetic and cultural ties with local Afro-Asiatic-speaking populations. By contrast, the Nuba/Noba today speak Kordofanian languages, which belong to the Niger-Congo family. They inhabit the Nuba Mountains/Kordofan Mountains/Nuba Hills to the south of the Nubians. The Noba/Nuba are thought to descend from the Nilotic-related inhabitants of ancient Nubia (the "black Nubians") because they share close physical, genetic and cultural ties with local Nilo-Saharan-speaking populations.)
 The researchers obtained mitochondrial sequences from eight X-Group skeletons buried at the Missiminia Necropolis, as well as from two Meroitic (including Late Meroitic) specimens and one Christian period individual interred at the site. They observed that the X-Group representatives carried mtDNA haplogroups that are nowadays associated with both Africa and Asia, including the X (twice), T1a, N, L3b, L3e, L2 and L1b clades. By contrast, both of the Meroitic samples belonged to the H2 lineage and the Christian era specimen was assigned to the W1 haplogroup, maternal clades that are of non-African origin. The scientists therefore postulate that there was in Nubia an "influx of sub-Saharan African ancestry after the Meroitic Period." This is indeed suggested by the presence of the L1b and L2 haplogroups in the X-Group/Post-Meroitic sample set. It is also supported by the fact that most modern Sudanese "Arabs" (Gaalien, Meseria, Arakien) carry mtDNA derivatives of the L macroclade (Hassan et al. (2009)). However, the L3 subclades are not necessarily indicative of Sub-Saharan introgression since the earliest occurrence of the L3 macrohaplogroup has, in fact, been found among specimens belonging to the Pre-Pottery Neolithic culture of the Levant (cf. Fernández et al. (2014)). Babalini et al. (2002) and Yatsishina et al. (2021) also each detected the L3 macroclade in, respectively, an ancient Libyan individual excavated in the Fezzan (dated to c. 3000 to 1500 BCE) and an ancient Egyptian mummy examined at the Kurchatov Institute. Additionally, Farrell et al. (2013) identified certain L3-rich areas in the Arabian Peninsula, where the local populations do not show any apparent Sub-Saharan genomic influence. Ergo, some of the L3 sublineages borne by the ancient X-Group individuals as well as many modern Afro-Asiatic and Nubian speakers in Africa could actually have been imported from the Middle East. (*N.B. For the latest evidence on the suggested non-African origin of the mtDNA macrohaplogroup L3, see Cabrera (2022) and Cabrera et al. (2017).)
The researchers obtained mitochondrial sequences from eight X-Group skeletons buried at the Missiminia Necropolis, as well as from two Meroitic (including Late Meroitic) specimens and one Christian period individual interred at the site. They observed that the X-Group representatives carried mtDNA haplogroups that are nowadays associated with both Africa and Asia, including the X (twice), T1a, N, L3b, L3e, L2 and L1b clades. By contrast, both of the Meroitic samples belonged to the H2 lineage and the Christian era specimen was assigned to the W1 haplogroup, maternal clades that are of non-African origin. The scientists therefore postulate that there was in Nubia an "influx of sub-Saharan African ancestry after the Meroitic Period." This is indeed suggested by the presence of the L1b and L2 haplogroups in the X-Group/Post-Meroitic sample set. It is also supported by the fact that most modern Sudanese "Arabs" (Gaalien, Meseria, Arakien) carry mtDNA derivatives of the L macroclade (Hassan et al. (2009)). However, the L3 subclades are not necessarily indicative of Sub-Saharan introgression since the earliest occurrence of the L3 macrohaplogroup has, in fact, been found among specimens belonging to the Pre-Pottery Neolithic culture of the Levant (cf. Fernández et al. (2014)). Babalini et al. (2002) and Yatsishina et al. (2021) also each detected the L3 macroclade in, respectively, an ancient Libyan individual excavated in the Fezzan (dated to c. 3000 to 1500 BCE) and an ancient Egyptian mummy examined at the Kurchatov Institute. Additionally, Farrell et al. (2013) identified certain L3-rich areas in the Arabian Peninsula, where the local populations do not show any apparent Sub-Saharan genomic influence. Ergo, some of the L3 sublineages borne by the ancient X-Group individuals as well as many modern Afro-Asiatic and Nubian speakers in Africa could actually have been imported from the Middle East. (*N.B. For the latest evidence on the suggested non-African origin of the mtDNA macrohaplogroup L3, see Cabrera (2022) and Cabrera et al. (2017).)
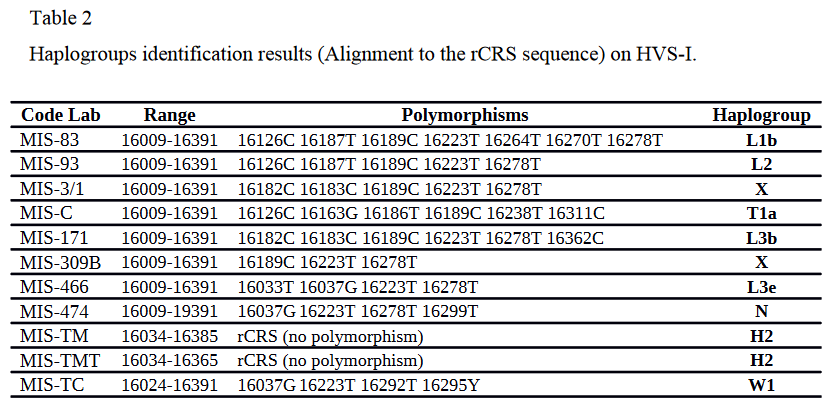
mtDNA haplogroups of ancient X-Group (MIS-83, MIS-93, MIS-3/1, MIS-C, MIS-171, MIS-309B, MIS-466, MIS-474), Meroitic (MIS-TM, MIS-TMT), and Christian (MIS-TC) period specimens buried at Missiminia, Upper Nubia (Cherifi and Amrani (2020)).
Now that we have both Y-DNA and mtDNA data on the X-Group/Post-Meroitic population, we can try and ascertain the prevailing direction of gene flow into this ancient community. More specifically, we can find out if:
1) The X-Group was originally a Nilo-Saharan-speaking population related to Nilotes, which later interbred with Afro-Asiatic speakers.
or
2) The X-Group was originally an Afro-Asiatic-speaking population related to Egyptians, which later interbred with Nilo-Saharan speakers and adopted from the latter the Nubian language.

mtDNA haplogroups of modern Nubians and Afro-Asiatic, Nilo-Saharan and Bantu-speaking populations in East Africa. 50%-60% of the Nubian and Afro-Asiatic-speaking individuals belong to maternal lineages derived from the Eurasian M and N macroclades; the remainder carry L(xM,N) haplogroups primarily consisting of the Pre-Pottery Neolithic-associated L3 clade (Non (2010)).
The uniparental markers would appear to favor scenario #2 since most of the X-Group individuals thus far analyzed (2 out of 3 samples or 66.67%) belong to paternal lineages derived from the YAP (DE) clade, like the preceding Meroites and most other neighboring Afro-Asiatic speakers. A minority (1 out of 3 samples or 33.33%) were assigned to haplogroup A, which is the modal patrilineal clade among the ancient and modern Nilotes of the Nile Valley (see Yousif and Eltayeb (2009)'s analysis above). If we assume that the L3 mtDNA sublineages that the X-Group population bears are of Middle Eastern extraction, this implies that most of the Post-Meroitic individuals had Afro-Asiatic-affiliated maternal lineages. This would, in turn, suggest that the Sub-Saharan influence primarily entered the X-Group population through assimilation of Nilotic men. In further support of this male-mediated admixture, Hrdy (1978), who analyzed the hair morphology of Meroitic and X-Group mummies buried at Semna South in Sudanese Nubia, reports that:
Although there is not a consistent statistically significant difference between the X-group and Meroitic samples, it is interesting that the X-group sample, especially the males, had higher curling variables, indicating more of an African element. [*N.B. Hrdy's African comparison sample consists of modern individuals from Kenya, Uganda, Mozambique and Zambia, who generally have similar hair texture as the Nilotes of the Nile Valley; this "East Africa" sample was first used in his 1973 study.]

Distribution of blond, brown, black and red pigmentation in hair samples from Meroitic, X-Group/Post-Meroitic and Christian era mummies buried at Semna, Sudanese Nubia. Most of the ancient specimens have brown hair. Also notice how reddish and blond hair are most common during the Meroitic period and gradually decline in the later X-Group/Post-Meroitic and Christian epochs, likely due to foreign contacts (Hrdy (1978)). (*N.B. Lazaridis et al. (2022) analyzed ancient Levantine individuals dating from the Bronze and Chalcolithic Ages and did not observe any examples of red-haired specimens. However, the scientists did find some ancient European samples with red hair. This suggests that the occurrence of red hair among the Meroites and ancient Egyptians is specifically attributable to intermingling with early European populations (cf. Supplementary Materials).)
The Meroites, for their part, show an even more frankly Egyptian uniparental marker profile. According to Yousif and Eltayeb (2009), 3 out of 4 or 75% of their Meroitic period samples culled from the Haraz Cemetery carry derivatives of the YAP/DE paternal haplogroup. Furthermore, Sirak et al. (2021) observed a prevalence of the Y125054 subclade of E1b1b-V12 among Christian-era Nubian individuals inhumed at the Kulubnarti site in Sudan. Since the medieval Kulubnarti specimens appear to descend from the Post-Meroitic or X-Group peoples that immediately preceded them, and the Post-Meroitic folks in turn seem to descend from the Meroites, it is likely that the YAP/DE derivatives which Yousif and Eltayeb found among their Meroitic cohort similarly comprise E1b1b sublineages. (*N.B. Trombetta et al. (2015) report that 74.5% of their modern southern Egyptian samples are haplogroup E1b1b-V12 carriers (cf. Supplementary Table 7). This points to close ties between Egyptians and the Christian-era, Post-Meroitic and Meroitic populations of Nubia.)
-

- Statue of the Meroitic Queen or "Candace" Amanirena (r. 40 BCE-10 BCE). Note the "Caucasoid" facial features and Berber-like oblique eye folds — immortalized via the Eye of Horus or wadjet symbol — which are so characteristic of ancient Egyptian art.
-

- Sculpture of the 18th Dynasty ancient Egyptian Treasurer Maya (right) and his wife, the Priestess Merit. The resemblance to the Meroitic statuary is conspicuous, notably in terms of the fine-features and almond-shaped eyes.
-

- Ancient Egyptian statue of Lady Sennuwy, wife of Djefaihapi, the provincial governor of Asyut (c. 1971-1926 BCE, 12th Dynasty). The bust was found in the tomb of a ruler of Kerma in Nubia. Notice again the similarity in style and features with the Meroitic sculpture.
-

- Modern statue of a Somali woman. The sculpture's resemblance to the Meroitic and ancient Egyptian figures is notable, and ultimately stems from shared ancestry between the Afro-Asiatic-speaking populations in the Horn of Africa and Nile Valley.
On the other hand, Cherifi and Amrani posit that the H2 mitochondrial clade, which both of their sampled Meroitic specimens belong to, was likely of Middle Eastern introduction. However, the H lineage also occurs among modern Nubians and contemporary Afro-Asiatic speakers in the Nile Valley and Horn of Africa (cf. Non (2010), Table 3-3). For example, Comas et al. (1999) note that a number of the Somali individuals they examined bore the Cambridge Reference Sequence (CRS), which belongs to the mtDNA haplogroup H. The lineage is particularly common among the Tuareg Berbers of the southern Maghreb, though here most individuals bear the H1 subclade (Ottoni et al. (2009)); the H clade is also widespread among the Asni, Bouhria and Figuig Berbers of Morocco (Coudray et al. (2009)). Among other ancient specimens, Breidenstein et al. (2018) indicate that all of the medieval Nubians they analysed at the El-Kurru site in Sudan carried Eurasian mtDNA lineages, with 2 of the 5 examined individuals (40%) bearing the H2 subhaplogroup. Similarly, Gad et al. (2020a) report that the 18th Dynasty ancient Egyptian Pharaoh Amenhotep III carries the H2b maternal clade. Drosou et al. (2020), moreover, divulge that the 25th Dynasty (c. 660 BCE) ancient Egyptian mummy Takabuti, an aristocrat from Upper Egypt, belongs to the H4a1 mtDNA sublineage. The scientists indicate that this is "a predominantly European haplogroup." Accordingly, this clade has been observed in a Neolithic Sardinian specimen (Marcus et al. (2020)). Rayo et al. (2022) likewise state that one of the canopic jars they analyzed, which contains the vital organs of a mummified ancient Egyptian individual, yielded the mitochondrial haplogroup H. The other container the researchers examined held viscera associated with the R0a1 clade, a maternal lineage that is today relatively common among Afro-Asiatic speakers in the Nile Valley, Horn of Africa and Arabian peninsula. (*N.B. Rayo et al. (2022) do not indicate any dates for their canopic jars, so it is unclear which period(s) these mitogenomes belong to.)
-
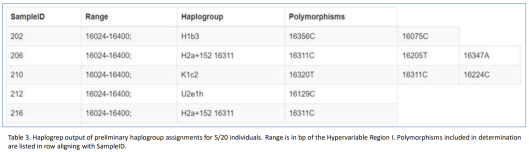
- mtDNA haplogroups carried by medieval Nubian individuals entombed at El-Kurru, Sudan. All of the examined specimens bear Eurasian maternal lineages, with H2 being the most frequently occurring clade (2 out of 5 individuals or 40%). This haplogroup has also been observed among earlier Nubian specimens dating from the Meroitic and Post-Meroitic/X-Group periods (Breidenstein et al. (2018)).
-
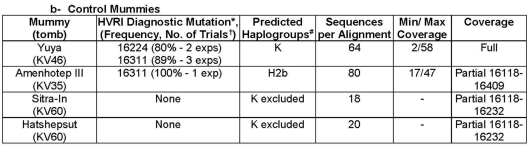
- mtDNA haplogroups borne by ancient Egyptian royal mummies. The 18th Dynasty Pharaoh Amenhotep III (b. 1411 BCE) was likewise found to belong to the H2 clade (Gad et al. (2020a)).
These findings only reinforce the Egyptian-like nature of the ancient Meroitic population — an aspect that was already suggested by artwork depicting Meroitic figures with a "Caucasoid" physiognomy, the Meroites' pyramid-building culture, their ancient Egyptian-derived writing system, and craniometric affinities (cf. Batrawi (1946); Mukherjee (1955); Billy (1981b); Rösing (1990)). As Breidenstein et al. cogently put it, "Nubian genetic makeup before Arab expansion is speculative, inferred from marker typing, or based on non-genetic markers, but thought to be closely related to Egyptians." (*N.B. Sirak et al. (2021), who analysed Christian-era individuals buried at Kulubnarti, similarly assert that "the rise (~300 BCE) and collapse (~350 CE) of the Meroitic Kingdom in Nubia provides a possible historical context for admixture between Egyptian peoples carrying West Eurasian-related ancestry and local Nubians." However, Sirak et al. argue that modern Nubians don't descend from the Christian-era Kulubnarti individuals (and, by extension, from the Meroites), but instead derive most of their West Eurasian ancestry from recent Arab Muslim settlers. This claim is rendered doubtful by the fact that a) the researchers utilized a Eurasian-admixed Dinka Nilote sample to arrive at their conclusions (cf. Skoglund et al. (2017); Scheinfeldt et al. (2019)), and b) Dobon et al. (2015), among other scientists, have observed that contemporary Nubians and Sudanese "Arabs" actually share most of their West Eurasian ancestry with Copts and other Afro-Asiatic speakers in Northeast Africa rather than with peninsular Arabs. Thus, it would make more sense to use ancient Egyptian samples, as opposed to Levantine or Arabian samples, as a proxy for the West Eurasian ancestry in modern Nubians; especially since, by Sirak et al.'s own admission, the ancient Egyptian specimens that have thus far been sequenced have just ~5% Sub-Saharan admixture (similar to the small amount of Eurasian admixture that has been detected in the much-used Mota sample) and are the best-fitting surrogate for the Kulubnarti individuals' West Eurasian ancestry.)

Craniometric analysis of ancient and modern populations from North Africa, the Horn of Africa, and the Middle East. Among the ancient samples, the predynastic Egyptian Badarians cluster nearest to the post-Neolithic populations of Nubia (C-Group, Kerma, Meroitic, X-Group, Philae) and the ancient Egyptians excavated at Deir el-Medineh, Upper Egypt. Among the recent samples, the Badarians cluster nearest to the Galla & Somali lumped sample (Rösing (1990)).
Hrdy (1978) likewise observed, with respect to the Meroitic and X-Group hair samples he examined, that:
In component I, which is heavily loaded on general curling variables and scale count, the total sample centroid was significantly different from European and African samples, though it was definitely more European than African.
-

- Wavy and curly hair strands belonging to ancient Egyptian mummies (Kurchatov institute).
-

- A soft-textured, wavy lock of hair belonging to a Kerma period individual excavated at Kadruka, Upper Nubia (northern Sudan). Ancient DNA analysis by Wang et al. (2022) has revealed that this specimen is genetically indistinguishable from the Cushites of the Early Pastoral Neolithic (see Genetic affinities of the Cushitic, Ethiosemitic and North Omotic-speaking populations of the Horn of Africa).
- Hair strands belonging to a Somali individual. Analysis of hair morphology has found that the Cushitic-speaking populations of Northeast Africa have a high proportion of individuals with soft-textured hair. Hair of this type is qualitatively the same as that observed among Meroitic specimens and ancient Egyptian mummies. This is consistent with a southward diffusion from the Nile Valley into the Horn of Afro-Asiatic-speaking settlers bearing West Eurasian ancestry.
As with the Meroites of Nubia, the anatomist Grafton Elliot Smith similarly remarks that most of the predynastic Egyptian specimens that he forensically analyzed — which number in the thousands, exponentially more than any other Egyptologist — had soft-textured wavy, straight or curly hair of a very dark brown or black hue:
The hair of the Proto-Egyptian was precisely similar to that of the brunet South European or Iberian people of the present day. It was a very dark brown or black colour, wavy or almost straight, and sometimes curly ; but it presented no resemblance whatever to the so-called "woolly" appearance and peppercorn-like arrangement of the Negro's hair.

Mummy of the ancient Egyptian Queen Tiye (1389-1338 BCE), "The Elder Lady", with her natural wavy red hair still attached. Forensic examination of ancient Egyptian specimens indicates that a significant minority had red hair. However, most predynastic Egyptian individuals possessed dark brown or black hair, like the related Meroites (cf. Elliot Smith (1911)).
-

- Ancient Egyptian sculptures of Queen Tiye wearing different royal headdresses. Archaeogenetic analysis of Tiye's mummy indicates that she bears the mtDNA haplogroup K, a European Neolithic-associated maternal lineage (Gad et al. (2020a); Gad et al. (2020b)). Furthermore, autosomal STR analysis suggests that Tiye and other members of the Amarna royal family carry a predominant non-African ancestry, with a close genetic affinity to populations in South Asia and Europe (LOP (2023)).

{kind=link}
- Colossal statue of Pharaoh Amenhotep III and his wife Queen Tiye (Egyptian Museum).
This also brings to the fore the question of the Meroitic language's origin. Is it an Afro-Asiatic idiom, as Kirsty Rowan (2006) contends? Or is it, as Claude Rilly (2010) argues, a Nilo-Saharan tongue, which we may presume an Egyptian-related, originally Afro-Asiatic-speaking population later adopted? Either hypothesis is plausible in view of the existing ancient DNA and morphological analyses.

A Meroitic statuette of a bound Noba ruler (50-1 BCE). The figurine has an engraving on it in the Meroitic language, which reads: qo qore Nob-o-l-o ("this one is the king of the Noba"). This inscription suggests that the Meroites were an ancestrally distinct people from the Kordofanian-speaking Noba. It, in turn, lends credence to the idea that either the Meroitic language belongs to the Afro-Asiatic family or the Meroites were originally Afro-Asiatic speakers who later adopted a Nilo-Saharan tongue (Donsmaps).

The Necropolis at Meroë has over 200 pyramids, exceeding in number those erected in ancient Egypt (Sacred Sites).
{kind=link}
In April 2020, Abigail Breidenstein of the Institute of Evolutionary Medicine at the University of Zürich and colleagues published an intriguing abstract on paleogenetics from Nubia. Although very brief, it nicely complements Yousif and Eltayeb's earlier work and Cherifi and Amrani's latest analysis. The researchers examined the mitogenomes of specimens dating from the Meroitic period through to the Christian era. They observed that most of the individuals belonged to Eurasian haplogroups (4 out of 6 samples or 66.7%), with the rest carrying maternal lineages derived from Sub-Saharan Africa. From the few details provided, it is uncertain what the identified haplogroups are or whether they include the Pre-Pottery Neolithic-associated L3 mtDNA clade. Nonetheless, it is interesting to note once more the close ties between the Meroitic, Post-Meroitic and Christian era populations of Nubia and the Afro-Asiatic speakers.
*Update #2
Using the Vahaduo Admixture JS program, genome analysis of Christian-era individuals excavated at the Kulubnarti site in Sudan indicates that they bear a predominant non-African ancestry (over 70%), consisting of majority West Eurasian elements (ancient Egyptian, European-related Steppe, and Levantine Natufian components) and a minority East Eurasian element (East Asian component). Furthermore, these ancient specimens have minor Sub-Saharan African admixture (under 27%) and a minute North African Iberomaurusian/Taforalt admixture (under 3%). The Cushites of the Pastoral Neolithic have the same ancestral composition, as do modern Cushitic, Ethiosemitic and North Omotic-speaking populations of the Horn region and Kababish "Arabs" from Sudan. Contemporary Muslim Egyptians and Libyans also share a similar ancestral makeup, but experienced some extra recent gene flow from western Asia. By contrast, Coptic Egyptians and northern Egyptians from Cairo and Mansoura appear to descend directly from earlier Dynastic period ancient Egyptians (as represented by the aristocrat Nakht-Ankh discussed above), with little apparent influences from other population sources. Maghrebi groups have an ancestral composition comparable to that of Libyans, with greater Iberomaurusian ancestry (~29% on average) as well as a significant Anatolian Neolithic admixture (~23%), especially in northern coastal areas opposite Iberia (refer to Genetic affinities of the Cushitic, Ethiosemitic and North Omotic-speaking populations of the Horn of Africa for the full details).

-
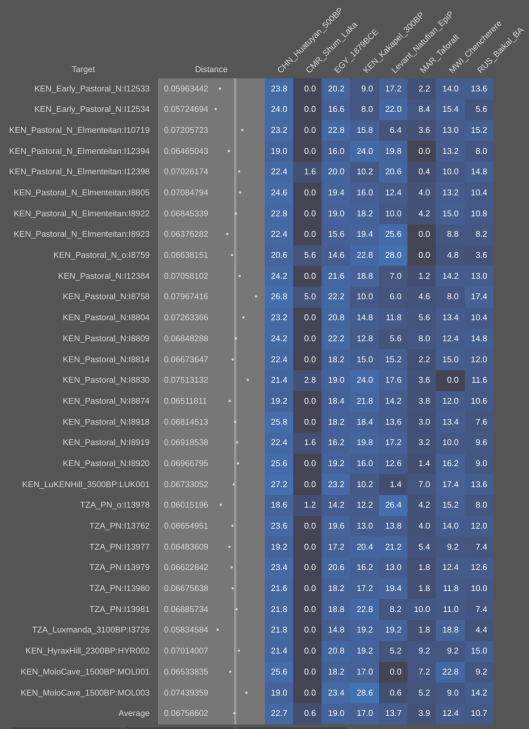
- Pastoral Neolithic
-
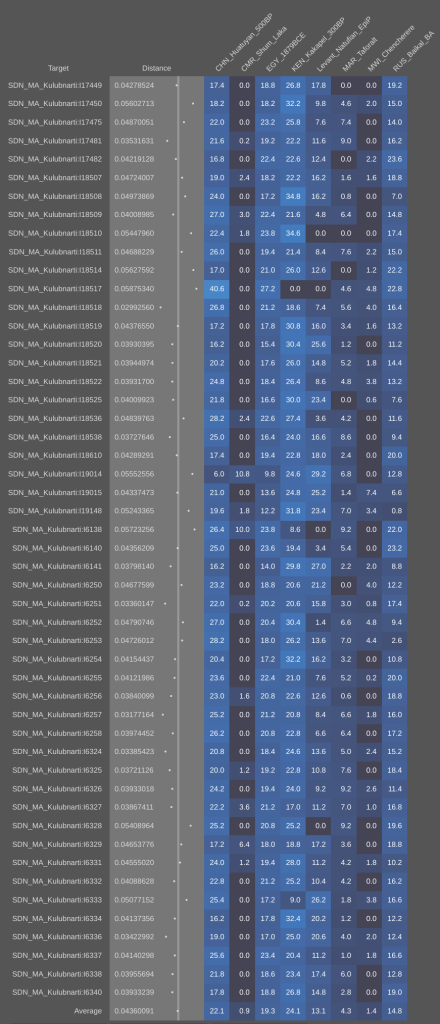
- Kulubnarti
-
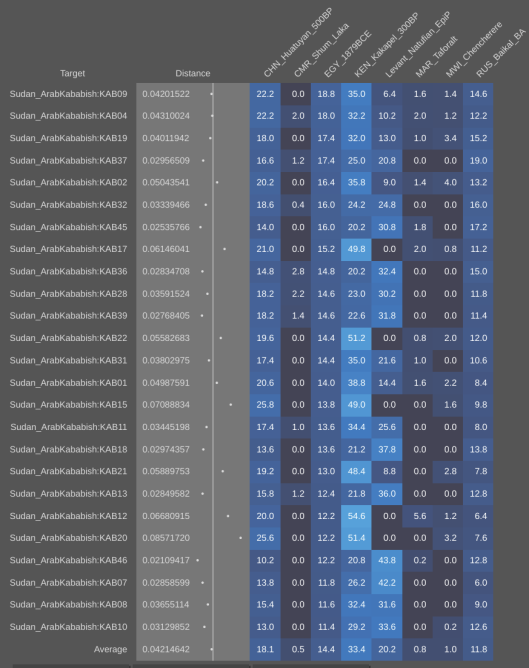
- Kababish "Arab" (Sudan)
-- Sent from my Linux system.
No comments:
Post a Comment